
Résumé d’un article de Malagoli, M., Sut, S., Kumar, G. et al. paru en juin 2022 dans la revue Chemical and Biological Technologies in Agriculture volume 9, 36.
Titre original : Variations of elements, pigments, amino acids and secondary metabolites in Vitis vinifera (L.) cv Garganega after 501 biodynamic treatment.
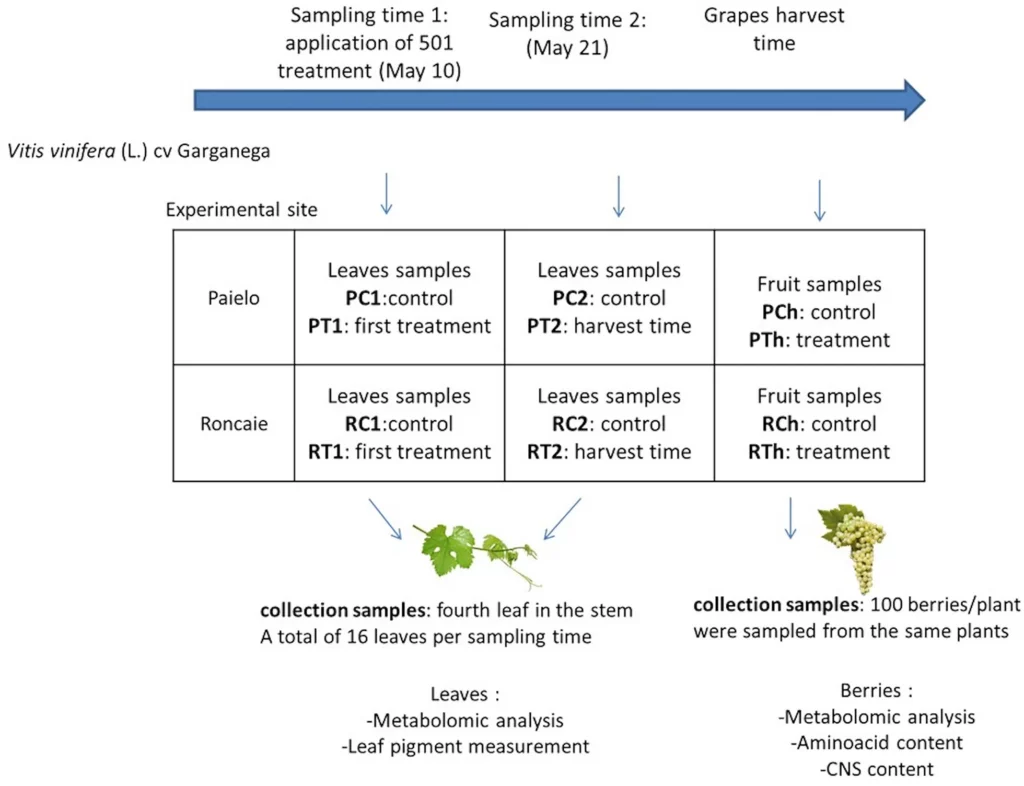
Présentation de l’essai
L’agriculture biodynamique se distingue de la culture biologique par l’utilisation de préparations spécifiques, appliquées sur les cultures ou le sol en très petites quantités. Ces préparations sont réputées stimuler le cycle des nutriments dans le sol, la photosynthèse et l’évolution optimale du compost, améliorant ainsi la qualité du sol et des cultures [2]. Les recherches sur la viticulture biodynamique ont révélé des similitudes avec le système biologique en ce qui concerne les caractéristiques du sol, la croissance et le rendement des plantes, l’utilisation des ressources et la biodiversité [22]. L’utilisation de préparations a eu une influence mineure sur la croissance et le rendement et n’a pas affecté la qualité finale des baies de raisin [23].
L’application de la silice de corne (préparation 501) sur la partie aérienne des plantes est l’une des recommandations données par Steiner pour mettre en œuvre l’agriculture biodynamique. Ce travail a pour but de mettre en évidence les effets de la pulvérisation de 501 sur les plantes de Vitis vinifera cv. Garganega.
Des échantillons de feuilles et de baies ont été prélevés sur des plantes cultivées dans deux vignobles, Roncaie (1ha) et Paiele (0,5 ha), situés 500m de distance sur la commune de Gambellara au nord-est de l’Italie. Les échantillons de feuilles ont été prélevés à deux moments, 6 heures après le traitement à la 501 (prélèvement 1, 10 mai) et 11 jours plus tard (prélèvement 2, 21 mai) afin d’élucider si le traitement à la 501 pouvait induire un changement immédiat dans le contenu des feuilles ou si un effet durable pouvait être détecté 11 jours après l’application. Des analyses ont également été effectuées sur des baies échantillonnées au moment de la récolte.
Pour évaluer les variations induites par l’application de 501, nous avons d’abord examiné la teneur en chlorophylle des feuilles, communément considérée comme un marqueur de l’état nutritionnel des plantes. L’absence de différence au niveau des pigments observée entre les feuilles des plantes témoins et celles des plantes traitées avec la 501 confirme les résultats d’une étude précédente sur des vignes traitées en biodynamie [9].
La métabolomique est appliquée avec succès en science végétale [24, 25], et les progrès techniques et la possibilité d’acquérir un grand nombre de données brutes, grâce à la spectrométrie de masse à haute résolution couplée à la chromatographie en phase liquide, permettent d’obtenir un profil phytochimique complet, ouvrant ainsi de nouvelles opportunités de recherche [26, 27, 28]. L’analyse statistique multivariée est un outil permettant d’étudier les altérations métaboliques dans des échantillons complexes comme les tissus végétaux. L’analyse en composantes principales (ACP) est l’une des techniques les plus utilisées dans l’analyse multivariée, qui vise à réduire un ensemble de données à ses principales composantes et à faire ressortir les similitudes. L’ACP a été utilisée dans plusieurs études pour distinguer l’origine variétale et/ou géographique des jus de raisin et des vins [29]. D’autres types de traitement des données, comme PLS-DA et sa version implémentée OPLS-DA, peuvent offrir l’avantage d’une interprétation plus facile des modèles. La plupart des dispositifs expérimentaux « omiques » visent à comparer des échantillons entre un groupe contrôle et un groupe test (par exemple, une maladie ou un traitement). L’objectif d’une telle analyse différentielle est donc de construire un modèle capable de distinguer les classes d’observations et de fournir une interprétation significative des différences observées [30].
Nous avons étudié les profils des métabolites dans les feuilles et les baies par analyse UPLC-QTOF-MS en utilisant une approche métabolomique non ciblée. Contrairement aux méthodes ciblées, où la plupart des métabolites présents dans la matrice sont ignorés, la métabolomique non ciblée vise à obtenir une couverture métabolique aussi large que possible sans supervision, y compris pour les composés inconnus. Par conséquent, les métabolites mesurés ne sont pas prédéfinis et le développement et la validation de la méthode suivent un schéma différent de celui de l’analyse ciblée [31].
Résultats
L’ACP pour les métabolites foliaires des deux vignobles a montré une classification uniquement due à la date d’échantillonnage, suggérant une variation limitée de la composition des feuilles par le traitement à la 501 par rapport aux changements causés par le temps. Cependant, une élaboration plus poussée par des méthodes supervisées a permis de détecter des changements dus au traitement, en évitant tout biais lié au moment et au lieu d’échantillonnage. Nos résultats sont, à notre connaissance, la première démonstration de changements métabolomiques liés aux applications biodynamiques de 501 sur les feuilles de vigne.
Voies métaboliques
Les changements de métabolites dans les feuilles des vignes de Paiele et de Roncaie ont indiqué que le métabolisme secondaire était principalement influencé 11 jours après le traitement biodynamique 501, ce qui permet d’exclure un effet à court terme (après 6 heures). La voie métabolique du shikimate menant à la formation de phénylpropanoïdes, de stilbénoïdes et de flavonoïdes semble être déclenchée dans les feuilles des plantes de Paiele traitées par la 501, tandis que l’augmentation des acides caftarique et caféique dans les vinges de Roncaie indique que la biosynthèse des phénylpropanoïdes est principalement influencée dans les plantes traitées. La forte augmentation des acides caféique et caftarique pourrait être contrebalancée par la baisse du niveau de resvératrol et d’épicatéchine dans les feuilles de Roncaie. La production de resvératrol dans la vigne est liée à des stimuli externes, comme une infection microbienne, une irradiation lumineuse, un stress hydrique, des éliciteurs ou des composés de signalisation [32]. Il est connu qu’une plus grande tolérance des plantes aux stress abiotiques est liée à une augmentation de la synthèse des polyphénols, tels que les acides phénoliques et les flavonoïdes [33]. Nos résultats suggèrent que l’application de la préparation 501 peut déclencher la biosynthèse d’antioxydants bénéfiques pour améliorer la tolérance au stress des vignes, bien que les différentes variations dans les niveaux de métabolites foliaires enregistrés dans les plantes des deux vignobles puissent être attribuables à un effet de site.
Acides aminés
Récemment, on a constaté que la teneur totale en acides aminés augmentait pendant la maturation dans les baies de raisin saines en biodynamie par rapport au traitement intégré [23]. Dans notre étude, la quantité totale d’acides aminés libres dans les baies n’a pas varié, bien que la 501 ait induit des variations significatives de certains acides aminés. Cependant, le manque de cohérence des changements entre les baies des deux vignobles conduit à exclure un effet commun de la 501 sur la biosynthèse des acides aminés, à l’exception de l’augmentation du niveau de cystéine. Il est connu que la quantité d’acides aminés individuels dans les baies peut varier en fonction de la variété, du lieu, de l’âge, des pratiques culturales et de la méthode d’analyse [34]. Dans une étude précédente, les analyses 1H-RMN des baies de Vitis vinifera L. cv. Sangiovese ont mis en évidence une accumulation plus importante de proline, de valine et d’isoleucine dans les baies biodynamiques que dans les baies biologiques [11]. Laghi et al. [14] ont comparé le vin rouge obtenu à partir de raisins biodynamiques et biologiques et ont conclu que la conduite du vignoble n’a entraîné que des modifications limitées sur la composition du vin. Les auteurs ont signalé des effets sur la proline, l’acide aspartique et la valine, les alcools et certains polyphénols.
Composés phénoliques et caroténoïdes
Les baies des plantes traitées avec la 501 dans les deux vignobles ont présenté un modèle superposable de modifications métaboliques, principalement identifiés comme des phénols et des caroténoïdes. Les changements métaboliques observés dans les baies différaient de ceux trouvés dans les tissus foliaires, ce qui peut être lié à la spécificité métabolique des deux organes de la plante. Les composés phénoliques sont importants dans le raisin en raison de leur fonction protectrice contre le stress environnemental et les infections fongiques [35]. Dans les plantes traitées avec la 501 sur les deux sites, le niveau des constituants phénoliques des baies a augmenté, mais seule la violaxanthine a augmenté de manière significative. Les caroténoïdes sont des photo-protecteurs bien connus dans les tissus des plantes et peuvent interférer pendant le processus de maturation du raisin [36]. Les baies répondent métaboliquement à la lumière en augmentant le niveau de composés comme les polyphénols qui ont des capacités antioxydantes directes et protègent les baies de la lumière solaire [37]. L’augmentation de l’épigallocatéchine, bien que seulement dans les baies de Roncaie, et de la violaxanthine peut être considérée comme un effet bénéfique du traitement biodynamique en termes de réponse accrue au stress oxydatif. Les polyphénols des baies de raisin sont extraits pendant la vinification et peuvent influencer la couleur ainsi que la perception sensorielle du vin [38]. De plus, les caroténoïdes du raisin peuvent jouer un rôle important en tant que précurseurs potentiels des composés aromatiques, comme par exemple les C13-norisoprénoïdes, qui sont responsables d’un impact sensoriel significatif dans les vins [39].
Un article précédent a étudié la composition phénolique des baies de raisin du cépage Pignoletto et du cépage Sangiovese, en comparant la gestion conventionnelle, biologique et biodynamique [40]. La catéchine et l’épigallocatéchine avaient une teneur comparable et la rutine a diminué dans le traitement biodynamique par rapport à la gestion conventionnelle et biologique des raisins de Pignoletto. Même Parpinello et al. (2019) n’ont mis en évidence aucune différence significative entre les vins biologiques et biodynamiques de Sangiovese, bien que ces derniers présentaient des concentrations plus élevées de malvidine-3-glucoside et de catéchine [10].
Nos résultats ont révélé que la catéchine et l’épigallocatéchine ont augmenté dans les raisins issus des parcelles de Paiele et Roncaie, ce qui suggère une tendance homogène du traitement biodynamique, indépendamment du lieu. Le traitement par la 501 a favorisé une augmentation générale des composés phénoliques, à l’exception du glucoside du kaempférol, dans les raisins des deux vignobles. Ces résultats sont en accord avec Reeve et al. (2005) qui ont rapporté une augmentation des polyphénols dans le vin obtenu à partir de raisins en culture biodynamique [21]. L’augmentation des polyphénols dans les raisins cultivés en biodynamie peut confirmer l’hypothèse d’une régulation positive des métabolites supposés induire une résistance au stress [21, 22]. Des niveaux plus élevés de flavonoïdes et d’anthocyanines, avec le potentiel antioxydant plus élevé qui en découle, ont été associés à une moindre vigueur de la plante, une caractéristique souvent signalée dans le cadre de la gestion biodynamique [41].
Globalement, nos données indiquent une stimulation des voies de biosynthèse des composés phénoliques dans les feuilles et les baies grâce à l’application de la 501.
Conclusion
La présente étude démontre que des approches complémentaires, combinant des données ciblées et non ciblées liées au métabolisme primaire et au métabolisme secondaire, peuvent offrir une nouvelle opportunité dans l’évaluation des effets complexes et multifactoriels des préparations biodynamiques en viticulture.
L’application de 501 sur des plantes de Vitis vinifera cv Garganega a favorisé des modifications du contenu des métabolites phénoliques dans les feuilles et les baies. Le niveau de la plupart des composés identifiés par des approches métabolomiques a augmenté dans les plantes traitées avec la préparation biodynamique 501. Une réponse commune au traitement à la 501 a été mise en évidence dans les constituants phénoliques des raisins, tandis que dans les feuilles, l’effet était détectable après 11 jours et dépendait du site de culture. Le métabolisme secondaire contribue à l’adaptation des plantes à l’environnement. La comparaison avec les résultats obtenus par d’autres études ne permet pas de suggérer une influence générale de l’application du 501 sur les feuilles et les baies de la vigne. Le nombre de viticulteurs qui s’intéressent à la culture en biodynamie est en augmentation dans le monde entier, étant donné le succès effectif de ce mode de culture et de la vinification. La présente étude peut être considérée comme un premier effort pour mieux comprendre les effets réels des préparations biodynamiques et pour informer les agriculteurs des connaissances scientifiques liées à la biodynamie. Des recherches supplémentaires sont assurément nécessaires pour explorer davantage le rôle de la silice de corne sur le métabolisme secondaire de la vigne.
Références
Voir l’article original.
Droits et autorisations
Accès libre. Cet article est soumis à une licence internationale Creative Commons Attribution 4.0, qui autorise l’utilisation, le partage, l’adaptation, la distribution et la reproduction sur n’importe quel support ou dans n’importe quel format, à condition de citer de manière appropriée le ou les auteurs originaux et la source, de fournir un lien vers la licence Creative Commons et d’indiquer si des modifications ont été apportées. Les images ou tout autre matériel tiers figurant dans cet article sont inclus dans la licence Creative Commons de l’article, sauf indication contraire dans une ligne de crédit vers le matériel. Si le matériel n’est pas inclus dans la licence Creative Commons de l’article et que l’utilisation que vous souhaitez en faire n’est pas autorisée par la réglementation ou dépasse l’utilisation autorisée, vous devrez obtenir l’autorisation directement auprès du détenteur des droits d’auteur. Pour consulter une copie de cette licence, visitez le site http://creativecommons.org/licenses/by/4.0/. La renonciation de la licence Creative Commons du domaine public (http://creativecommons.org/publicdomain/zero/1.0/) s’applique aux données mises à disposition dans cet article, sauf indication contraire dans la ligne de crédit des données.